Síntesis de proteínas
Las proteínas, por su tamaño, no pueden atravesar la
membrana plasmática de la célula; por eso, existe en su
interior un mecanismo que las construye (síntesis) según
las necesidades que tenga en ese momento la célula.
La síntesis de proteínas o traducción tiene lugar en
los ribosomas del citoplasma celular. Los aminoácidos
son transportados por el RNA de transferencia (RNAt),
específi co para cada uno de ellos, y llevados hasta el RNA
mensajero (RNAm), donde se aparean el codón de éste y
el anticodón del RNA de transferencia, por complementariedad
de bases, y de esta manera se sitúan en la posición
que les corresponde.
Cuando termina la síntesis de una proteína, el RNAm
queda libre y puede leerse de nuevo. De hecho, es muy frecuente
que antes que fi nalice una proteína, se inicia la lectura
para otra, con lo cual una misma molécula de RNAm
es utilizada por varios ribosomas simultáneamente.
Este proceso es de fundamental importancia, ya que
básicamente todas las características que presenta la
célula (fenotipo) se regulan por la suma de sus actividades
enzimáticas. En pocas palabras, todo lo que la célula
es y puede realizar depende de la acción enzimática
específi ca. Como casi todas las enzimas son proteínas,
la morfología y funcionamiento celular depende del tipo
de proteína que la célula debe armar. Con el transcurso
de la evolución, todos los organismos se aseguraron
que la información correspondiente para sintetizar sus
enzimas específi cas esté presente en sus células y en su
descendencia.
CARACTERÍSTICAS DEL RNAT
La síntesis del RNAt se realiza a través de la catálisis de
la polimerasa del RNA III tal y como se vio en el capítulo
de transcripción. Éste se encuentra disperso por todo el
citoplasma; es el más pequeño de los tres tipos de RNA
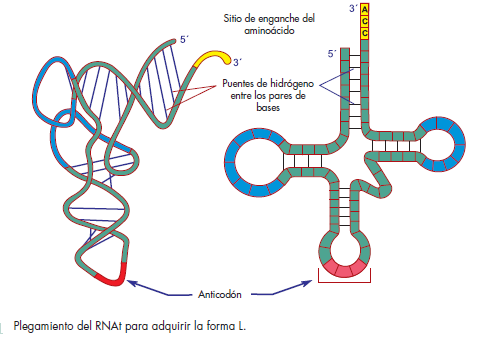
y su estructura tiene forma de hoja de trébol. Los RNAt
se estructuran por alrededor de 80 nucleótidos con pesos
moleculares de cerca de 25 000 daltones (fi g. 6-1).
Todos los RNAt tienen pG en el extremo 5′ y pCpCpA
en el extremo 3′. El extremo 3′ se conoce como brazo del
aminoácido o también brazo de unión al aminoácido o
“aceptor”. El correspondiente brazo del anticodón contiene
el triplete anticodón, el cual reconoce el codón del
RNAm y se relaciona con éste por medio de formación
de puentes de hidrógeno, siguiendo las reglas de complementariedad
de las bases.
Cada tipo de RNAt lleva antepuesto el nombre del
aminoácido que transporta. Por ejemplo, leucinil-RNAt
para la leucina, lisinil-RNAt para la lisina, fenilalanil-
RNAt para la fenilalanina, metionil-RNAt para la metionina,
etcétera. Sin embargo, para poder efectuar esta unión
y reconocimiento específi co del RNAt con su respectivo
aminoácido, se necesita la participación de una enzima

AMINOACILSINTETASA
Es la enzima que cataliza la activación y la unión del aminoácido
correcto al RNAt correcto. Son doblemente específi
cas por reconocimiento molecular:
1. Para cada aminoácido: reconocen propiedades de carga,
hidrofobicidad y tamaño.
2. Para cada RNAt correspondiente: interactúan específi
camente con el brazo aceptor y con el brazo del
anticodón (fi g. 6-2).
3. Conocen e interpretan el código genético.
Así, son capaces de corregir errores, pues contienen
sitios especiales de “revisión”; si el aminoácido es incorrecto,
se hidrolizan del RNAt y tienen alta fi delidad.
Existen 20 aminoacilsintetasas de RNAt diferentes,
cada una específi ca para reconocer a un aminoácido y al
RNAt compatible con él. Ambos reconocimientos permiten
que cada uno de los 31 tipos de RNAt se una a sólo
uno de los 20 aminoácidos utilizados durante la síntesis
proteínica. Ello es posible porque cada aminoacilsintetasa
de RNAt identifi ca al RNAt por el anticodón, la parte
más específi ca del RNAt.
ACTIVACIÓN Y UNIÓN
DEL AMINOÁCIDO AL RNAT
Los aminoácidos se activan por medio de las aminoacilsintetasas
específi cas y de ATP, antes de unir los aminoácidos
a su RNAt específi co.
Los aminoácidos, una vez activados, forman el complejo
aminoaciladenilato monofosfatado, liberando el
pirofosfato, producto secundario del ATP (fi g. 6-3). La
misma enzima aminoacilsintetasa localiza al RNAt específi
co para el aminoácido correspondiente y da lugar a
la formación del aminoacil-RNAt, liberándose la enzima
para reiniciar otro ciclo con otro aminoácido similar.

MADURACIÓN Y/O PREPARACIÓN
DEL RNAT PARA UNIRSE
AL RIBOSOMA
El trabajo de los RNAt consiste en tomar del citosol los
aminoácidos y conducirlos al ribosoma en el orden marcado
por los nucleótidos del RNAm, que son los moldes
del sistema.
Así, la función básica de los RNAt es alinear a los aminoácidos
siguiendo el orden de los codones para poder
cumplir con sus funciones; los RNAt adquieren una forma
característica semejante a un trébol de cuatro hojas. Los
cuatro brazos se generan por la presencia en los RNAt de
secuencias de tres a cinco pares de nucleótidos complementarios,
los cuales se aparean entre sí como los
nucleótidos de las dos cadenas del ácido desoxirribonucleico
(fi g. 6-5).
En la punta de uno de los brazos confl uyen los extremos
5′ y 3′ del RNAt. El extremo 3′ es más largo, de modo
que sobresale el trinucleótido CCA que fue incorporado
durante el procesamiento postranscripcional del RNAt.
Este brazo se llama aceptor, porque a él se liga el aminoácido,
que se une a la A del CCA.
Los tres brazos restantes poseen en sus extremos
secuencias de 7 a 8 nucleótidos no apareados, con forma
de asas, cuyas denominaciones derivan de los nucleótidos
que las caracterizan. Una de ellas contiene el triplete
de nucleótidos del anticodón, por lo que su composición
varía en cada tipo de RNAt. Otra, en virtud de que contiene
dihidrouridinas (D), se denomina asa D. La tercera
se conoce como asa T, por el trinucleótido TΨC que la
identifi ca. La letra T simboliza a la ribotimidina y la Ψ a
la seudouridina.
Entre el asa T y el anticodón, existe un asa adicional
llamada variable, porque su longitud difi ere en los distintos
RNA de transferencia.
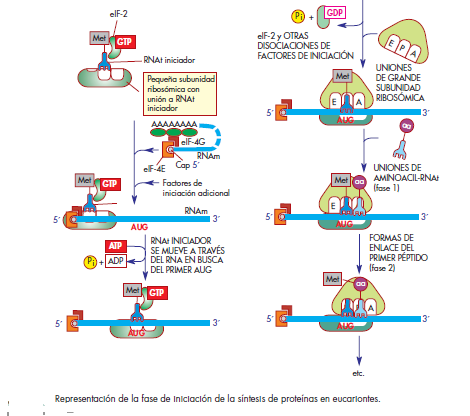
INICIACIÓN DE LA SÍNTESIS
DE PROTEÍNAS
Procariontes
Se comienza con la subunidad menor sola. IF-1 se une
a la base del sitio A para forzar que el primer fMet-RNAt
entre en el sitio P. IF-3 tiene una doble función, ya que se
le necesita para estabilizar la subunidad 30S y para que
el RNAm interaccione con dicha subunidad. IF-2 (como
otros muchos factores de traducción) es del tipo de proteínas
G que sirve para depositar el aminoacil-RNAt
(fMet-RNAt en este caso) en el ribosoma.
.
